
Serge‐Ely Dibakou
Preliminary assessment of gastrointestinal parasites of the sun-tailed monkey (Allochrocebus solatus) in a semi-free-ranging colony
Dibakou, Serge‐Ely; Ngoubangoye, Barthélémy; Boundenga, Larson; Ntie, Stephan; Moussadji, Cyr; Tsoumbou, Thierry‐Audrey; Setchell, Joanna M.
Authors
Barthélémy Ngoubangoye
Larson Boundenga
Stephan Ntie
Cyr Moussadji
Thierry‐Audrey Tsoumbou

Professor Jo Setchell joanna.setchell@durham.ac.uk
Professor
Abstract
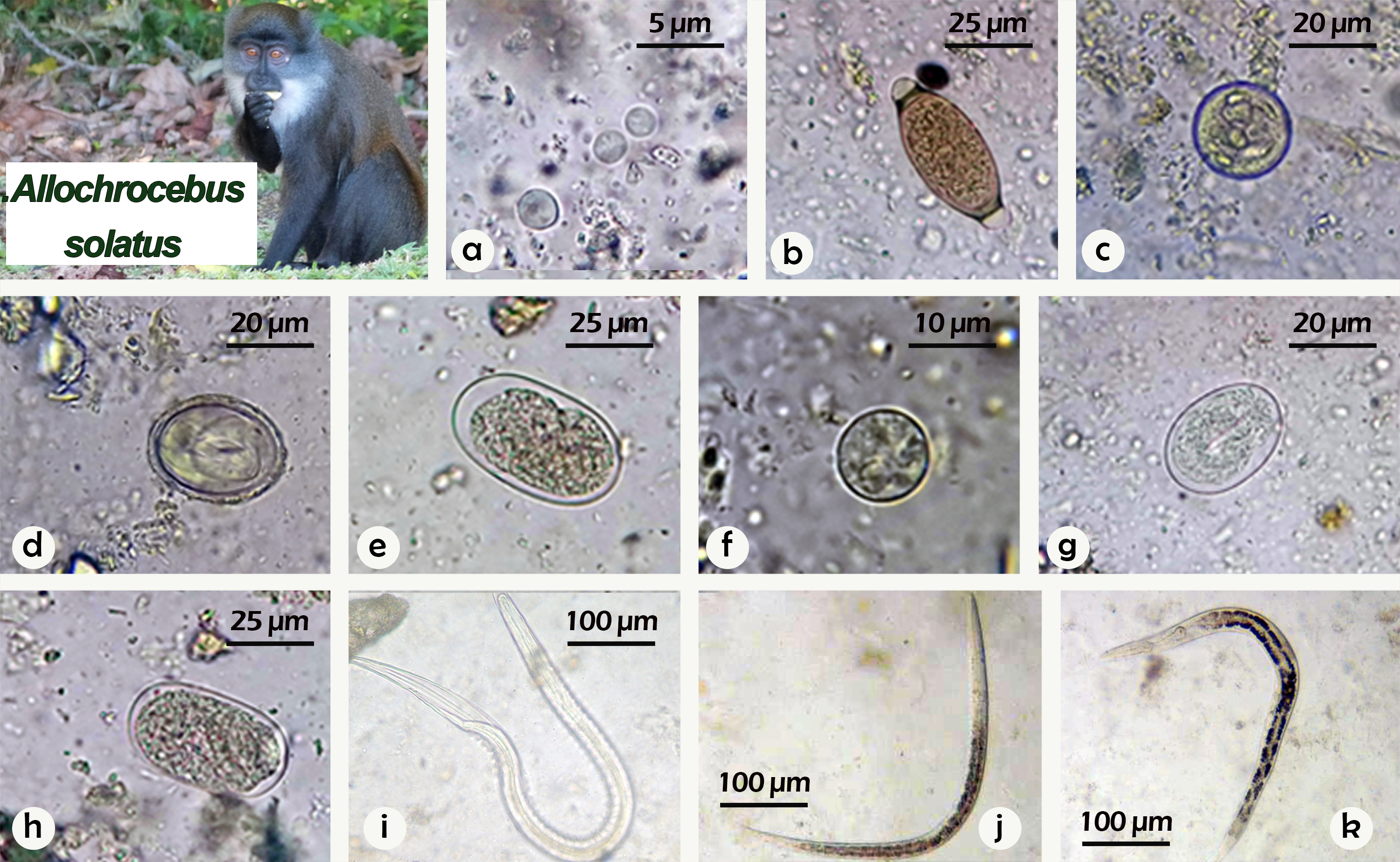
Background: The occurrence of gastrointestinal parasites in the sun-tailed monkey (Allochrocebus solatus) at the CIRMF primatology center is unknown. We, therefore, assessed the presence and richness (number of different parasite taxa) of gastrointestinal parasites in a semi-free-ranging colony of A. solatus. Methods: A total of 46 fecal samples were screened using a modified McMaster technique for fecal egg counts. Results: In the 46 samples collected, seven taxa of gastrointestinal parasites, including protozoa and nematodes were identified. The most prevalent parasite was strongyles parasites (98%), followed by Trichuris spp. (72%), Strongyloides spp. (67%) and Entamoeba coli (65%). Balantioides coli (33%), Endolimax nana (25%), and Spirurid eggs (26%) were only found in a minority of the animals. Conclusions: This study contributes new host records of gastrointestinal parasites in semi-free-ranging A. solatus and highlights the need to investigate the health of this species and implement proper precautions in the management of this colony.
Citation
Dibakou, S., Ngoubangoye, B., Boundenga, L., Ntie, S., Moussadji, C., Tsoumbou, T., & Setchell, J. M. (2022). Preliminary assessment of gastrointestinal parasites of the sun-tailed monkey (Allochrocebus solatus) in a semi-free-ranging colony. Journal of Medical Primatology, 51(3), 127-133. https://doi.org/10.1111/jmp.12581
| Journal Article Type | Article |
|---|---|
| Acceptance Date | Mar 15, 2022 |
| Online Publication Date | Mar 26, 2022 |
| Publication Date | 2022-06 |
| Deposit Date | Jun 16, 2022 |
| Publicly Available Date | Mar 26, 2023 |
| Journal | Journal of Medical Primatology |
| Print ISSN | 0047-2565 |
| Electronic ISSN | 1600-0684 |
| Publisher | Wiley |
| Peer Reviewed | Peer Reviewed |
| Volume | 51 |
| Issue | 3 |
| Pages | 127-133 |
| DOI | https://doi.org/10.1111/jmp.12581 |
Files
Accepted Journal Article
(214 Kb)
PDF
Copyright Statement
This is the peer reviewed version of the following article: Dibakou, Serge‐Ely, Ngoubangoye, Barthélémy, Boundenga, Larson, Ntie, Stephan, Moussadji, Cyr, Tsoumbou, Thierry‐Audrey & Setchell, Joanna M. (2022). Preliminary assessment of gastrointestinal parasites of the sun-tailed monkey (Allochrocebus solatus) in a semi-free-ranging colony. Journal of Medical Primatology 51(3): 127-133, which has been published in final form at https://doi.org/10.1111/jmp.12581. This article may be used for non-commercial purposes in accordance with Wiley Terms and Conditions for Use of Self-Archived Versions. This article may not be enhanced, enriched or otherwise transformed into a derivative work, without express permission from Wiley or by statutory rights under applicable legislation. Copyright notices must not be removed, obscured or modified. The article must be linked to Wiley’s version of record on Wiley Online Library and any embedding, framing or otherwise making available the article or pages thereof by third parties from platforms, services and websites other than Wiley Online Library must be prohibited.
Accepted Journal Article (Figure 1)
(2.3 Mb)
Image
Copyright Statement
Figure 1
You might also like
Chemical cues of identity and reproductive status in Japanese macaques
(2022)
Journal Article
Downloadable Citations
About Durham Research Online (DRO)
Administrator e-mail: dro.admin@durham.ac.uk
This application uses the following open-source libraries:
SheetJS Community Edition
Apache License Version 2.0 (http://www.apache.org/licenses/)
PDF.js
Apache License Version 2.0 (http://www.apache.org/licenses/)
Font Awesome
SIL OFL 1.1 (http://scripts.sil.org/OFL)
MIT License (http://opensource.org/licenses/mit-license.html)
CC BY 3.0 ( http://creativecommons.org/licenses/by/3.0/)
Powered by Worktribe © 2024
Advanced Search